Open Access | Review
This work is licensed under a Creative Commons Attribution-ShareAlike 4.0 International License.
Role of microRNAs in skin aging and its potential therapeutic interventions
* Corresponding author: Durairaj Sekar
Mailing address: RNA Biology Lab, Saveetha Dental College and Hospitals, Saveetha Institute of Medical and Technical Sciences,
Saveetha University, Chennai, India.
Email: duraimku@gmail.com
* Corresponding author: Deusdedit Tusubira
Mailing address: Department of Biochemistry, Mbarara University of Science and Technology, Mbarara, Uganda, P O Box 1410, Mbarara, Uganda.
Email: dtusubira@must.ac.ug
The article belongs to the Special Issue: Metabolic function, energetics and aging
Received: 23 April 2024 / Revised: 09 May 2024 / Accepted: 03 June 2024 / Published: 27 June 2024
DOI: 10.31491/APT.2024.06.144
Abstract
The skin, the largest and most important organ of the body, serves diverse functions such as protection from the outside, temperature regulation, etc. It is composed of 3 layers: hypodermis, dermis, and epidermis. As the skin ages, it undergoes morphological and anatomical changes. Wrinkle formation and sagging are the main physical changes visible on the skin. Aging is caused by extrinsic and intrinsic factors that affect the morphology and function of the skin. Intrinsic aging is driven by biological and genetic factors, while extrinsic aging is influenced by environmental factors. Recent findings show the contribution of miRNAs in skin aging, particularly in replicative senescence. They regulate the expression of genes post-transcriptionally and control functions like collagen modulation, elastin synthesis, etc. This review explores the role of miRNAs in skin aging and details their role in cellular functions, including collagen synthesis. This review also describes the types and characteristics of skin aging, structural alteration in different layers of skin, and therapeutic interventions of miRNA in skin aging. Knowledge of miRNA involvement in aging will help to introduce new therapies in antiaging and skin care technology. The intricate relationship between environmental and genetic factors that cause skin aging is discussed, emphasizing the role that miRNAs play in this complex process. In addition, potential therapeutic interventions are described, highlighting the need for deeper knowledge of the molecular mechanisms of skin aging for effective, innovative future research.
Keywords
Skin aging, miRNA, collagen, cellular senescence, therapeutic interventions, skin structure
Introduction
As the largest and most diverse organ in the body, the skin performs a variety of functions, including regulating body temperature, shielding the body from external agents and infections, and resisting moisture. The entire skin layer serves as a barrier to protect the body from the elements [1]. It consists of 3 distinct layers: the hypodermis, dermis, and epidermis. The epidermis is primarily composed of keratinocytes. It is also composed of melanocytes and Merkel cells. The second layer, the dermis, helps with blood and nutrient circulation. It consists of blood vessels, connective tissue, nerves, etc. The hypodermis, the bottom layer of the skin, stores energy and connects the skin to bones and muscles. It is composed of connective tissue and adipose tissue [1, 2]. Aging is an inevitable phenomenon that leads to a gradual loss of tissue integrity. Figure 1 shows the structure of normal and aged skin.
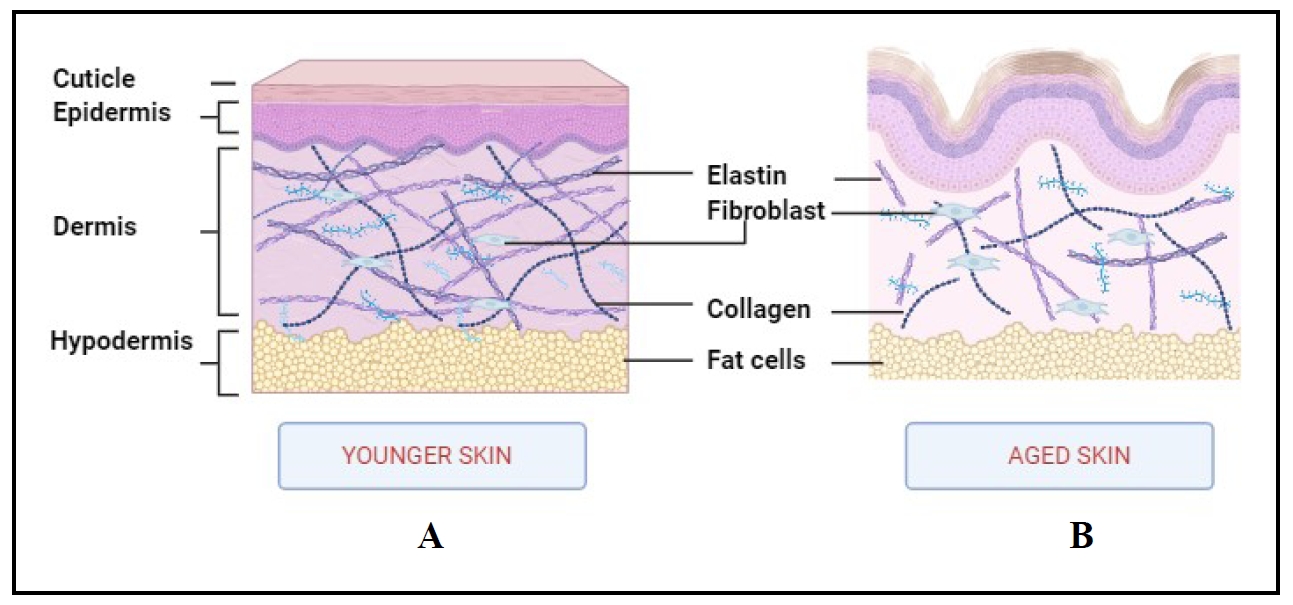
Figure 1. The structure of aged skin in comparison to normal young skin. (A) Normal skin contains elastin,collagen, fibroblasts, and fat cells in different skin layers in a connected manner. (B) Aged skin with disrupted elastin collagen, fibroblasts, and fat cells in skin layers. During aging, the synthesis of collagen and elastin decreases and their network breaks down, leading to wrinkling and sagging of the skin.
The most apparent manifestation of this skin aging is characterized by various changes, including the development of wrinkles,
sagging, and pigmented spots [3]. The skin is the most visible organ in the body and all
alterations, including aging, are easily observed. Aging is a broader concept encompassing overall alterations in the appearance
of skin and dermal tissues, along with their functions.
It is interesting to note that cellular senescence is a concept that describes the condition in which individual cells experience
permanent growth arrest. Hayflick and Moorhead were the first to define aging at the cellular level by demonstrating the limited
ability of human primary fibroblasts to divide. The replication process causes telomeres to become unable to maintain their length,
which results in the so-called Hayflick limit. As a result, cells lose their ability to proliferate and enter an irreversible
cell cycle arrest known as cellular senescence [2, 4].
Cellular senescence is one of several factors that lead to skin aging.
Skin aging is a more complex phenomenon involving both morphological and functional changes in the skin over time. Moreover, the
aging phenomenon affects the skin in various ways, including thinning of the epidermis and a remarkable decrease in the turnover
rate. The integrity of the skin and its physiological features are also affected. Although men and women age differently, the
appearance of their skin also varies, but the internal and external factors are the same
[5, 6]. This natural progression involves a
complex interaction between intrinsic and extrinsic factors, which is why it is classified as intrinsic and extrinsic aging
[4, 6]. Genetic and biological components are
considered intrinsic or internal variables, whereas environmental exposures and lifestyle choices are extrinsic or external
factors. The effects of intrinsic aging on the skin are similar to those on other internal organs. It is also known as
chronological aging because time is another factor that causes it. In addition, the two main signs of natural aging are the
flattening of cells at the epidermal-dermal junction and the thinning of the dermis, which is a layer of skin. Moreover, elastin
and collagen in the skin break down over time, reducing the integrity and stability of the skin
[7]. In contrast, extrinsic aging, also known as photoaging, is induced by sun exposure
and manifests itself in exposed areas of the skin, such as the backs of the arms and the face
[8]. It is characterized by visible signs such as dark spots, wrinkles, and a rough,
leathery skin texture. Additionally, reduced elasticity, abnormal coloration, graying hair, and hair loss are other results of
photoaging [9].
Although skin aging is a normal aspect of life, the external indicators of aging can have a major impact on how someone feels
about themselves. Therefore, it is essential to understand the cosmetic elements. In addition to these cosmetic consequences,
skin aging also has functional consequences. With age, changes in the skin's structure can compromise its protective functions.
To detect abnormal growth and maintain homeostasis, the skin relies on molecular signals due to its diverse functions and cell
types [10]. These pathways contribute to morphological and functional changes in the
skin. However, a common hallmark of aging is the up- and downregulation of protein expression associated with the electron
transport chain. Whether this is a cause or a consequence of aging remains controversial
[11]. To understand the genetic aspects of aging, several studies have been conducted
on protein and gene expression in various aging model systems. An emerging focus in these investigations is on small noncoding
RNAs, particularly microRNAs, which act as potent post-transcriptional regulators [12].
Over the past decade, research on miRNAs has revealed their role in all cellular processes. These small regulatory molecules play
an integral role in fundamental cellular activities such as cell proliferation, differentiation, aging, etc.
[13]. They also play a crucial role as biomarkers in various diseases.
MicroRNAs (miRNAs) are non-coding RNAs found in all organisms and plants. They are approximately 25 nucleotides in length. These
molecules are involved in the posttranscriptional regulation of gene expression by binding to their target
[14]. Recently, miRNAs have been discovered to be significant regulators of aging and
senescence. These short RNAs play an important role in modulating the expression of mRNA targets, leading to translational
repression or degradation of mRNA. This promotes protein degradation or prevents translation
[7]. There are several miRNAs that are commonly involved in many skin activities,
including collagen modulation, keratinocyte differentiation, etc. Altered expression of miRNAs prevents proliferation of
cells, affects elastin production, imbalances skin cell turnover, increases oxidative stress, etc.
[2, 10]. Moreover, miRNAs have become common
regulators of a number of skin disorders and may serve as new biomarkers or therapeutic targets for etiology or treatment. In
short, miRNAs have a variety of functions in skin biology and their intricate roles make them more potential candidates for
further studies and research in the field of dermatology and skin therapy [2]. In the
present review, our objective is to focus on the contribution of miRNAs in skin aging. The primary objective of this review is to
deliver extensive knowledge in the field of skin aging and the role of miRNAs in the aging process. This understanding of miRNA
involvement in aging provides the basis for developing innovative therapies and pioneering in a new era of anti-aging interventions
and skin care technology.
Skin structure
The skin is the largest part of the body that provides protection from the external environment. It accounts for 8 to 15% of total adult human body weight and occupies 1.8 m2 of surface area [11, 15]. It is also a complex and dynamic organ with multiple functions. In addition, the skin maintains the homeostasis of the body by preventing the loss of electrolytes, proteins, and fluids, along with temperature regulation [1, 16]. The epidermis, dermis, and hypodermis are three distinct layers that make up the skin and are functionally connected layers [17]. The structure of the skin is illustrated in Figure 2.
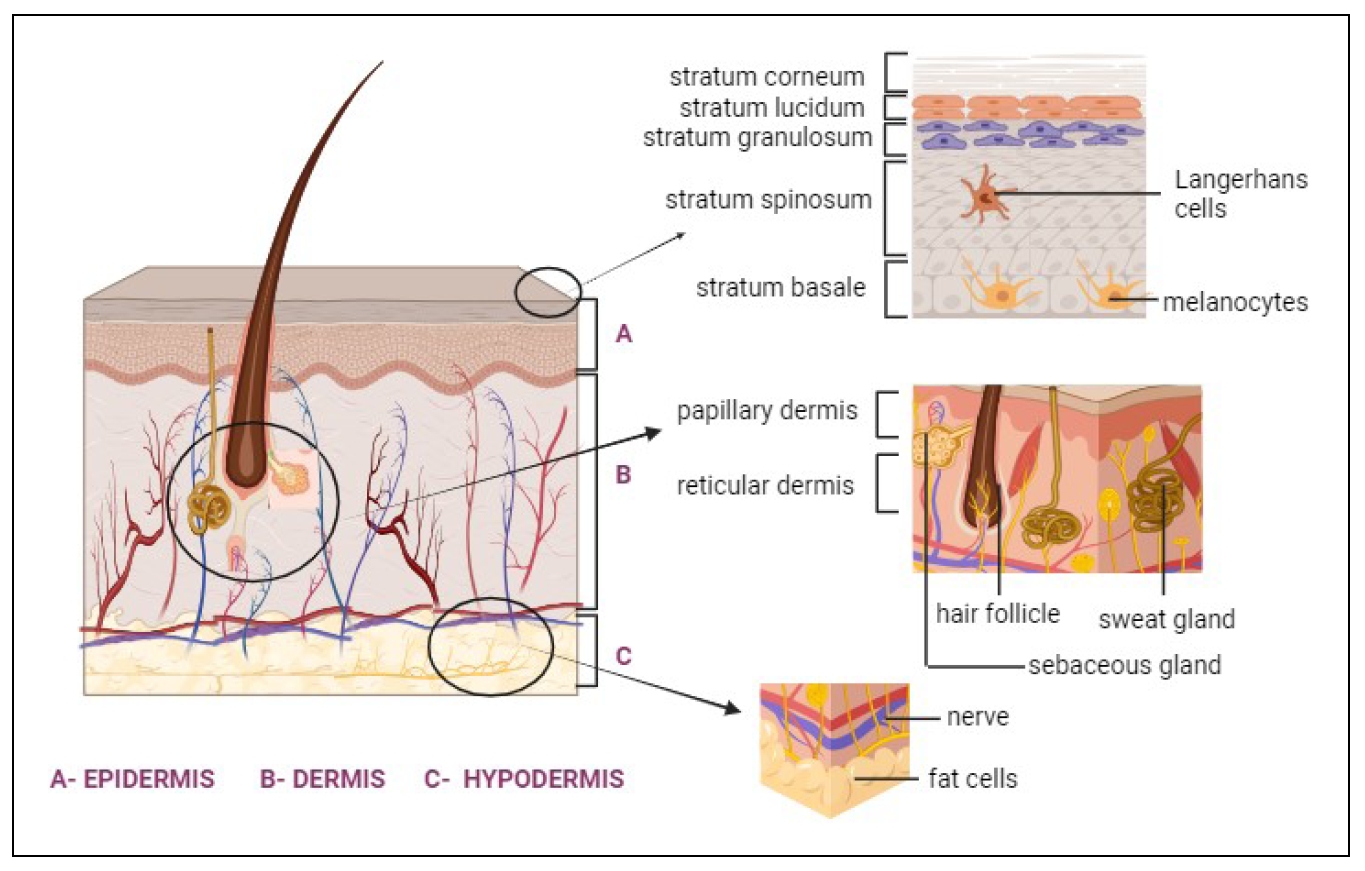
Figure 2. The structure of the skin. (A) EPIDERMIS is the outer layer of skin that comprises the Stratum corneum, Stratum lucidum, Stratum granulosum,Stratum spinosum, and Stratum basale. (B) DERMIS is the second layer below the epidermis. It consists of the Papillary dermis and the Reticular dermis. This layer is characterized by hair follicles, sweat glands, and sebaceous glands. (C) HYPODERMIS, which is the deepest layer, contains nerve and fat cells.
The outermost layer of the skin, derived from the ectoderm, is called the epidermis. It protects the body from the external
environment, including ultraviolet (UV) protection, thermoregulation, and immune protection. Keratinocytes, melanocytes,
inflammatory cells, Langerhans cells and neuroendocrine cells are the main cell types of the epidermis. Among these, keratinocytes
cover most of the epidermis, and they're interconnected by tight junctions [11].
They produce keratin, which contributes to the skin's waterproofing properties. Melanocytes synthesize the pigment melanin, which
provides color and UV protection [17, 18]. The
epidermis is arranged in several layers, including the stratum spinosum, stratum basalis, stratum granulosum, and stratum
corneum (Figure 2A). The outer layer of the epidermis is called the stratum corneum, which
is consists of flattened keratinocytes that provide protection from external factors. The stratum basale is the innermost layer of
the epidermis, where new cells are formed. As they move toward the surface, the cells eventually become flattened
[4]. In a recent study, keratin fibers stiffened in naturally aged skin and water movement
in the stratum corneum also decreased [19, 20].
The number of melanocytes and Langerhans cells decreased, leading to epidermal thinning
[19].
Beneath the epidermis is the dermis. The dermis is mainly involved in nutrient supply to the cells of the skin. It is a connective
tissue layer composed of collagen and elastin fibers that are interconnected. This layer is rich in lymph, nerves, hair follicles,
and sweat glands [2, 11]. It is also populated
with lymphocytes, macrophages, dendritic cells, etc. Apart from nutrient supply, it also provides oxygen and detects
touch and pain [1]. Dermis cells are mostly presented by fibroblasts and are predominantly
involved in extracellular matrix (ECM) components. The ECM is composed of collagen and glycosaminoglycan embedded with mast cells,
neural cells, and endothelial cells [21]. The dermis is divided into two layers: the
reticular and papillary dermis (Figure 2B), which work together to perform all functions.
The papillary dermis connects the epidermis and blood vessels, while the reticular dermis provides strength to the skin
[22]. The density and thickness of dermal collagen decreases in chronologically aged
skin. Although photoaging leads to a decrease in the number and size of blood vessels, it can induce angiogenesis
[19, 23].
The hypodermis, called the subcutaneous layer, is located just below the dermis and consists of loose areolar connective tissue
with abundant adipose tissue and less collagen content. It is the deepest layer of the skin
(Figure 2C). This layer enables cutaneous mobility. Metabolic energy is stored in the
adipose tissue of the hypodermis and it also provides thermal insulation [24]. This layer
contributes to the insulation of the body and offers a protective cushion for internal organs. Vitamin D is also produced in this
layer [1]. The hypodermis of aged skin exhibits reduced ability to both maintain
protective barriers and recover from damage [4,
25]. Aging also leads to decreased cell proliferation in the basal layer, causing the
hypodermis to thin and reducing contact between the dermis and epidermis. Consequently, this reduces the nutrient supply to the
epidermis and weakens the renewal capacity of cells [4].
Approximately 30,000 dermal cells perish every minute, emphasizing the continuous renewal processes inherent to the skin.
Successful skin development and regular physiological functions hinge on the coordinated interplay of genetic networks and
regulatory factors [26]. The skin maintains a constant self-renewal mechanism to
replenish injured or old cells in spite of constant exposure to environmental pollutions and facilitate tissue repair. However,
when its regenerative ability decreases with age, it can trigger cellular senescence, paving the way for the onset of various
dermatologic conditions [27].
Skin aging—types and characteristics
The appearance and functionality of the skin will gradually change due to numerous biological and physiological processes
leading to aging [28]. Skin wrinkling, dry skin, skin thinning and sagging are the main
characteristics [3]. In addition, it causes age spots, hyperpigmentation, decreased
elasticity, and other issues. The skin, which serves as the body's barrier, is constantly exposed to a variety of external and
internal stimuli, which causes a progressive loss of regenerative potential [4]. Along
with structural and functional alterations in extracellular matrix skin components such as collagen, elastin, and others, aging
processes also bring about phenotypic changes in cutaneous cells [29]. Skin aging occurs
in two forms: intrinsic and extrinsic. Natural aging, another term for intrinsic aging, is mainly caused by genetic factors,
whereas extrinsic aging is influenced by environmental factors such as sunlight [9,
13]. Apart from that, apoptosis, DNA damage and telomere shortening are some of the
aspects that cause skin aging.
Intrinsic aging is the term for a natural physiological process that causes wrinkling in dry skin
[9, 30]. Histologically, a flat epidermal-dermal
interface and the absence of dermal papillae are characteristics of skin that is intrinsically aged. However, normal epidermal
differentiation and cellular polarity appear to be preserved [13,
31]. Collagen synthesis is reduced, as is the ability to heal wounds. Moreover, atrophy
of the dermis may occur due to a reduced repair capability, which will not be able to restore collagen fibers that have been
degraded by this process [8, 32]. The number of
blood vessels is also decreased which causes a decreased blood supply to cells. Apart from genetic factors, other elements that
drive intrinsic aging are hormones and time. It is an interesting fact that the telomere, the terminal region of the eukaryotic
chromosome, significantly influences intrinsic aging. Telomeres contain a short sequence of nucleotides, TTAGGG. Repeated cycles
of replication ultimately lead to the loss of protection at the end of the chromosome (particularly this sequence), making it
susceptible to end-to-end fusions. This condition is incompatible with regular cell functioning. Most cells can divide up to 60 or
70 times over their lifetimes before entering senescence, a state in which they are still viable but unable to divide
[32]. Oxidative stress also contributes to skin aging, eventually causing the
accumulation of oxidative damage to proteins, lipids, and other constituents of cells. It's connected to a gradual reduction in
antioxidant capability and an increase in the formation of reactive oxygen species (ROS). Inflammation, which causes ROS production
as well as activation of pro-inflammatory cytokines [33]. Another element contributing
to skin aging over time is a modification in the levels of growth factors and apoptosis. Moreover, hormones like melatonin have
been observed to decrease as well. At the same time, signaling molecules also play a vital role in aging. Particularly, certain
signaling molecules became more abundant with age, whereas others, such as chemokines, declined and caused the destruction of
various skin functions [34, 35]. Histological
alteration observed in the skin includes changed stratum corneum permeability, decreased water loss that is attributed to a
hardened and thickened stratum corneum, and changed lipid composition causes greater cellular cohesion. In addition, the number of
fibroblasts and mast cells in the dermis also decreased [19].
Extrinsic aging is mainly affected by skin exposure to environmental factors such as sunlight and pollution. Lifestyle also
contributes to photoaging. These substances degrade the collagen and elastin fibers of the skin
[29]. Paul Gerson Unna observed in their study that there was a transformation in the
skin of sailors, particularly in areas exposed to the sun, leading to premature aging. Studies have shown that UV radiation is
the key element in extensive aging, which accounts for 80% of facial aging [36]. The
process of UV-induced skin aging is complex and can be initiated by multiple signaling pathways, such as telomere DNA destruction,
protein oxidation, mitochondrial damage, and other signaling initiated by receptors [37].
In addition, in the late 19th century, Harry Daniell found that smokers appeared older than non-smokers. Thus, alcohol and smoke
also contribute to extrinsic aging [38-40].
The histological signature of photoaging is characterized by dermal elastosis, marked by the presence of thickened, intertwined,
and finally granular elastic structures. In contrast to intrinsic aging, photoaged skin forms thick layers with irregular
pigmentation, large wrinkles, and elastosis [41]. Compared to chronologically aged skin,
photoaged skin has a thicker epidermal layer. Furthermore, UV exposure may increase the number of keratinocytes and melanocytes
[19, 42]. The prevalence of pathologically
altered elastic fibers, sometimes referred to as "solar elastosis", is the most prominent histological characteristic of
photoaging. The activation of matrix metalloproteinases (MMPs) is the main cause of the destruction of elastin and collagen
structures. Furthermore, lymphocytes, mast cells, eosinophils, and other inflammatory cells are elevated in extrinsic aging
[19]. However, all skin layers are affected by aging and show changes in their roles
and morphology [43].
Pathways involved in aging
Aging consists of several interconnected processes that are driven by genetic programming, continuous external inputs, and
internal metabolic reactions. All organs and systems are impacted by aging, although at different rates
[11, 44]. With aging, the morphological and
physiological functions of the skin, like all other organs, gradually deteriorate. As a result of the progressive diminishing of
cellular constituents, mechanical protection is weakened. This slows down the immune response, which leads to an imbalance in
thermoregulation [45]. Increased levels of matrix metalloproteinases (MMPs) and
matrix-degrading enzymes, released primarily by epidermal keratinocytes and dermal fibroblasts, cause a decrease in extracellular
matrix (ECM) formation in the dermis, which results in the morphologic manifestation [46].
These MMPs are involved in the degradation of collagen fibers, a characteristic feature of intrinsic aging, and the partial
degradation of elastin fibers, a process associated with extrinsic aging. Imbalances in oxidative stress and inflammatory
processes further intensify the activity of MMPs, which will also contribute to skin aging [30].
Although the two types of aging are distinct, it appears that identical molecular pathways underlie both forms of skin aging. In
fact, the breakdown of the extracellular matrix by upregulated MMPs and the production of ROS are shared characteristics of skin
aging in both types. ROS are free oxygen radicals and other oxygen-derived molecules that are highly reactive. With aging, ROS
accumulate and lead to the inactivation of protein tyrosine phosphatases (PTPs or RPTPs), thereby activating receptor tyrosine
kinases (RTKs). These RTKs can also be activated by other factors such as inflammatory cytokines and growth factors. Moreover,
RTK activation is not uniformly increased in aged skin; some RTKs may decrease with aging. Activated RTKs undergo phosphorylation,
triggering downstream signaling pathways. This phosphorylation cascade activates three families of mitogen-activated protein
kinases (MAPK), such as extracellular signal-regulated kinases (ERK), c-Jun N-terminal kinase (JNK) and p38MAPK. Subsequently, the
transcription factor AP-1 (activator protein-1) is activated downstream of these MAPK pathways. This activation leads to the
expression of various MMPs, including MMP-1, MMP-3, and MMP-9, while simultaneously inhibiting the expression of procollagen-1.
Basically, ROS-induced signaling prompts a cascade of molecular reactions that activate pathways involved in the breakdown of the
ECM and inhibit procollagen-1 expression. Inhibition of procollagen-1 expression disrupts the regular synthesis of collagen, which
causes the loss of structural and functional integrity of dermal tissues. Ultimately, it causes problems related to elasticity and
thus aging [19, 47,
48]. For instance, a study has shown that the epidermal growth factor receptor (EGFR)
exhibits decreased expression and phosphorylation of downstream signaling pathways in the epidermis. This decrease in EGFR activity
suggests that RTK regulation in aging skin is complex and varies among different RTKs [49].
The molecular mechanisms of ROS involved in skin aging compared to normal mechanisms are illustrated in
Figure 3 and Figure 4. Skin function and
morphology were also declined by the accumulation of senescent cells. Although the basic mechanisms are still being elucidated,
there is mounting evidence for the existence of diseasecausing pathways leading to cutaneous aging. Extrinsic and intrinsic aging
may occur simultaneously or concurrently. Genetic and epigenetic mechanisms control all these processes. Regulation of non-coding
RNAs such as miRNA, long ncRNA, etc., modification of histones and methylation of DNA are some of the mechanisms
involved [11]. Meanwhile, miRNAs remain one of the most mysterious aspects in the study
of aging biology, despite all the available data. The expression of certain miRNAs may serve as a biological indicator of aging,
including photoaging, natural aging, and age-related disorders [3].
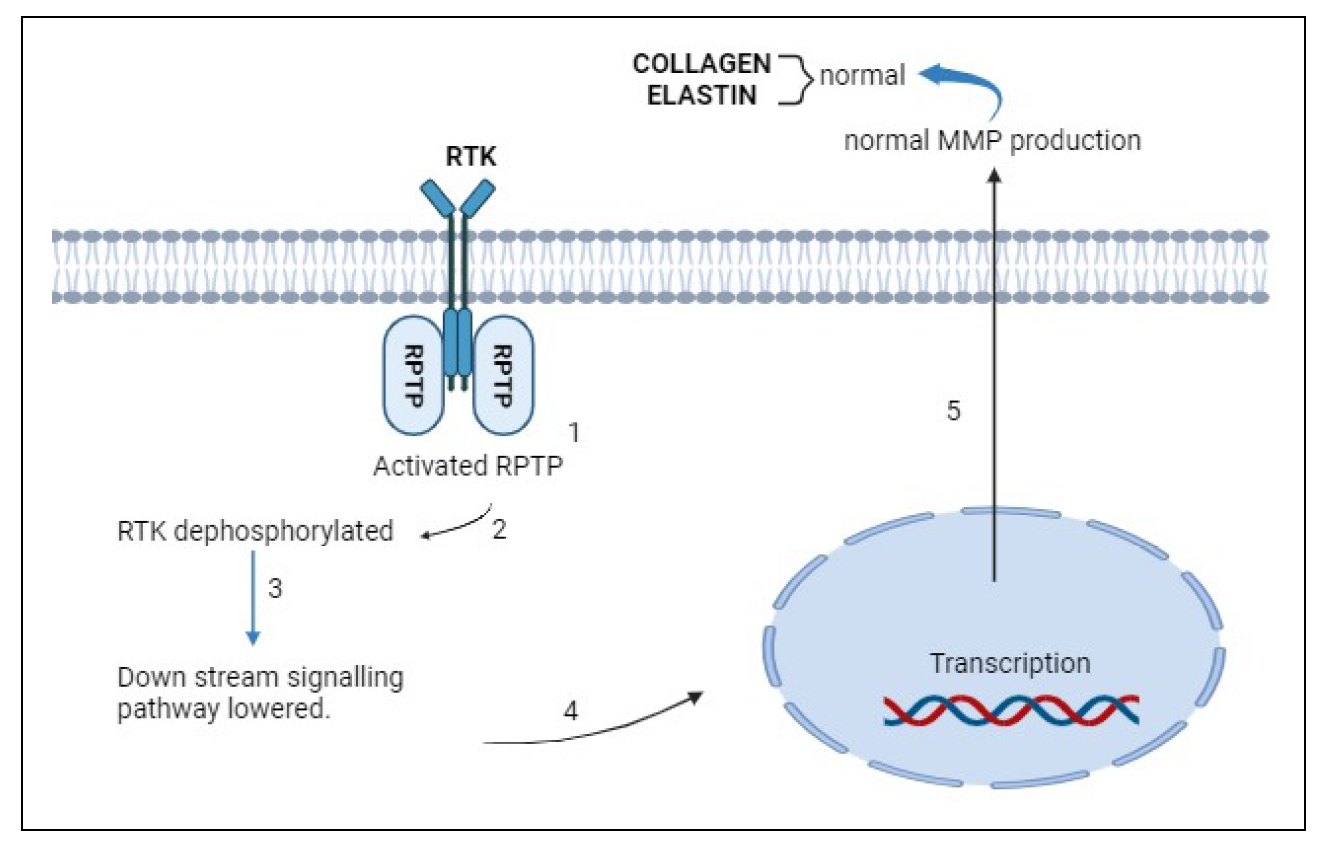
Figure 3. Formation of collagen and elastin in normal skin. (1) Under normal conditions, in the absence of ligands, receptor protein tyrosine phosphatase (RPTP) is activated. (2) It inhibits receptor tyrosine kinase (RTK) activity on the cell membrance by binding to it. (3) This dephosphorylated RTK downregulates the signaling pathway, leading to (4) transcription and (5) production of collagen and elastin.
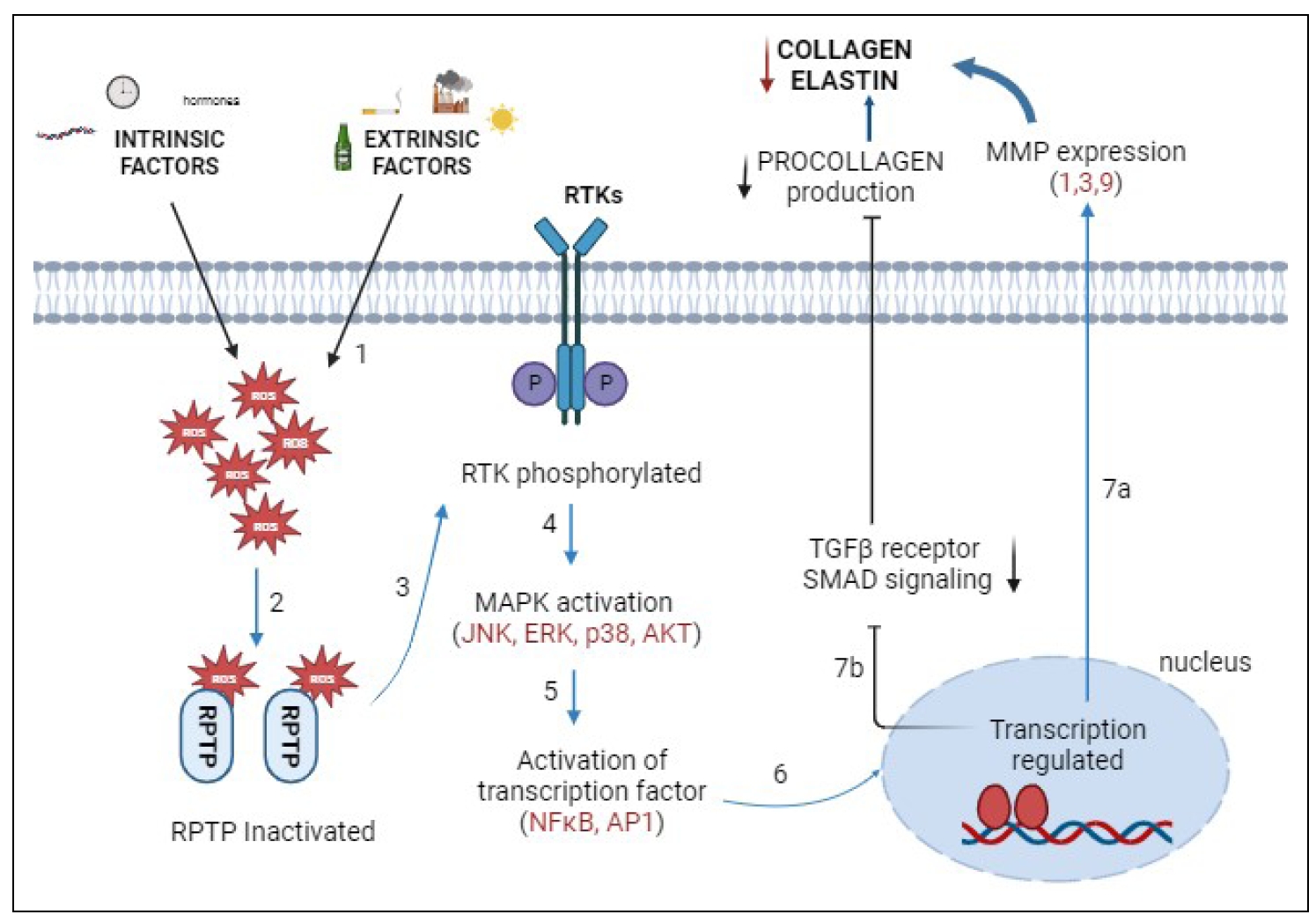
Figure 4. The pathway under the influence of intrinsic and extrinsic factors. (1) Intrinsic and extrinsic factors lead to the production of reactive oxygen species (ROS), which accumulate in the cell. (2) ROS inhibit RPTP by binding to the catalytic site. (3) RTK is phosphorylated and activated. (4) Activation of mitogen-activated protein kinases (MAPK) triggers other downstream signaling pathways, including extracellular signal-regulated kinases (ERK), c-Jun N-terminal kinase (JNK), protein kinase B (AKT), and p38. (5) Nuclear factor κB and transcription factor activator protein-1 (AP-1) are then activated. (6) This regulates gene expression. Altered gene expression inhibits collagen synthesis in two ways. (7a) The expression of MMPs (MMP-1, 3, 9) is increased. Collagen production is decreased. (7b) Inhibition of TGF-β and SMAD signaling pathways. Decrease the synthesis of procollagen 1, which reduces collagen and elastin.
Interaction of the somatotrophic system and miRNAs in skin aging
The somatotrophic system, which encompasses the somatotrophic axis and extrapituitary circuits, plays a crucial role in skin aging, especially in the context of congenital isolated growth hormone receptor deficiency (IGHD). IGHD, caused by a mutation in the GH-releasing hormone receptor (GHRH) gene, results in significantly reduced levels of GH and IGF-1, which are known for their cell proliferation and differentiation properties [50, 51]. Beyond the hormonal aspect, miRNAs have emerged as important regulators of metabolic changes associated with healthy aging. Our study hypothesized that GH deficiency in humans alters the abundance of circulating miRNAs, with a subset overlapping those found in GHdeficient mice [52, 53]. Among the significantly regulated miRNAs, hsa-miR-31, hsa-miR-146b, hsa-miR-100, hsa-miR-181b, hsa-miR-195 were of particular interest for their roles in aging-related pathways. In vitro assays confirmed that these miRNAs regulate the expression of age-related genes such as mTOR, AKT, NFκB, and IRS1, which are implicated in skin aging [52]. For instance, miR-181-5p was upregulated approximately sevenfold in IGHD humans, with an even more pronounced effect in older individuals. This miRNA inhibits cell proliferation, migration, invasion and tumorigenesis by targeting IGF-1R and its downstream signaling pathways. Additionally, miR-181b-5p expression is increased in human senescent keratinocytes, indicating its tissue-specific properties and potential contribution to skin aging [52, 54]. These findings highlight the complex interaction between the somatotrophic system and miRNAs in resulting skin aging. This will provide new insights into research and enhancing skin health.
Role of microRNAs in general aging process
Aging is a complex phenomenon characterized by progressive degradation and reduced repair capacity of tissues and organs, leading to age-related pathological disorders. miRNAs regulate gene expression by repressing translation and play a key role in aging [11, 55]. Key miRNAs such as miR-34a and miR-29 promote cellular senescence, while miR146-a and miR-21 modulate inflammation [56]. Recent research has focused on miRNAs as biomarkers of aging in humans.
Role of microRNAs in skin aging and therapeutic interventions
Short, single-stranded, non-coding RNAs are called microRNAs. They are about 25 nucleotides in length. These miRNAs regulate
gene expression [14]. Several miRNAs have been found in both plants and mammals. It was
first discovered in Caenorhabditis elegans, which is found in most eukaryotes, including humans. It is interesting to
note that most studies show that many protein-coding genes are regulated by these miRNAs at the translational and transcriptional
level. It is estimated that miRNAs account for about 1–5% of the human genome and control at least 30% of genes that code for
proteins [55, 57-59].
However, the findings of Boehm and Slack reveal the role of miRNAs that regulate the age of C. elegans and establish the
significance of miRNA in the skin aging process [60]. Furthermore, most miRNAs are
synthesized from DNA sequences to become primary miRNAs, or pri-miRNAs, which are then processed to become precursor miRNAs, or
pre-miRNAs, and ultimately mature miRNAs. miRNAs often decrease expression by interacting with the 3′ UTR of target mRNAs
[23]. However, there are also reports of miRNA interaction with other sites, including
the 5′ UTR, gene promoters, and the UTR at 5' region. Meanwhile, research has shown that miRNAs regulate the rate of translation
and transcription by shuttling between various cellular compartments [61,
62]. Many biological processes depend on these regulators. Moreover, it is necessary for
the normal growth in animals. Recent findings also revealed that miRNAs play a key role in regulating cutaneous aging and
senescence. Many of the miRNAs were up- or down-regulated with aging at the tissue or organism level
[11, 63]. In the context of the aging pathway
outlined before, miRNA has the capacity to regulate various molecules. It provides additional control over the signaling cascade
linked with ROS-induced aging. It is interesting to note that the first investigation of miRNAs associated with aging in human
epidermal keratinocytes revealed a set of regulated miRNAs [11].
miRNAs have the ability to regulate the levels of ROS either directly or indirectly. Moreover, it can regulate RPTPs, TGF-β and
other proteins associated with redox homeostasis, as well as proteins like RTKs, AKT, MAPKs, etc., thereby regulating the
downstream signaling cascade. Redox homeostasis refers to the balance between the production and elimination of ROS in the cell.
By targeting the mRNAs of these constituents, they can regulate ERK, JNK, etc. When miRNAs downregulate the expression
of PTPs and other proteins, it will result in the accumulation of ROS, which leads to the breakdown of the ECM and the inhibition
of procollagen 1. This will ultimately lead to the reduced production of collagen and the loss of structural and functional
integrity of dermal tissues, which is a hallmark of aging [64].
Figure 5 depicts the involvement of miRNAs in the regulation of gene expression in skin
biology at the post-transcriptional level (in various pathways).
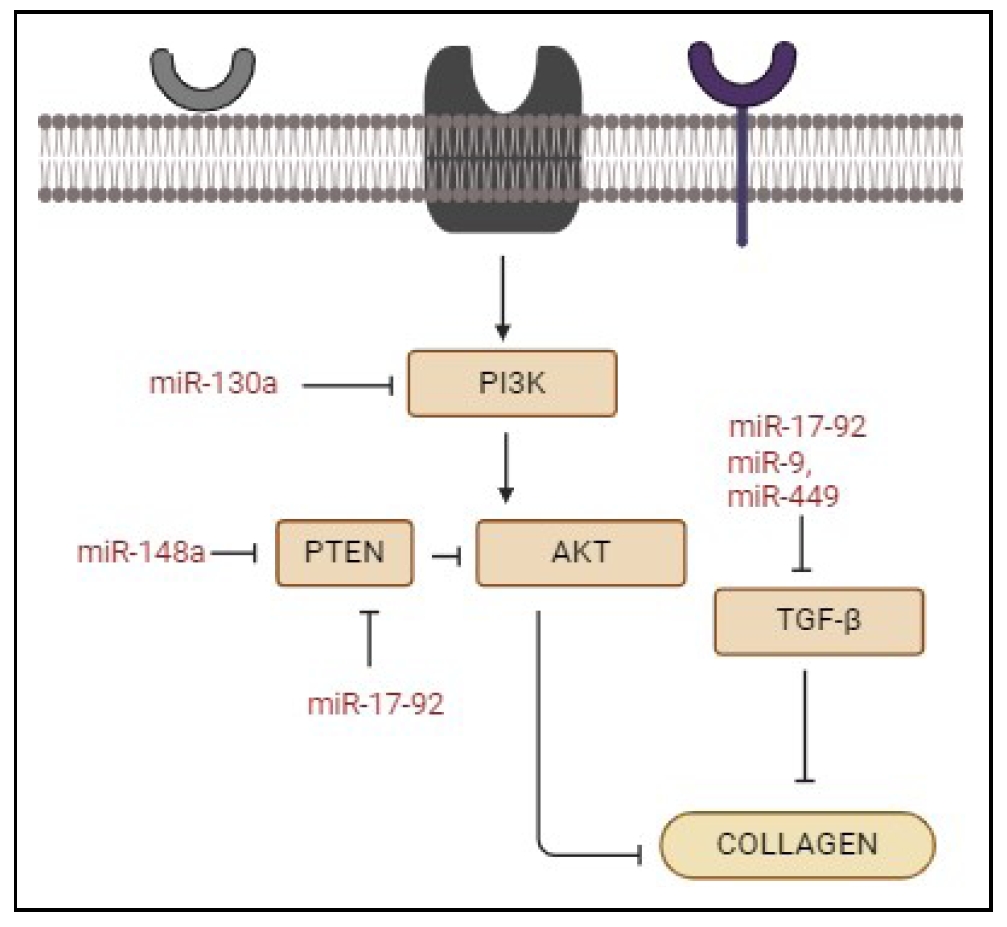
Figure 5. The role of different miRNAs in various signaling pathways leading to downregulation of collagen synthesis. PI3K, AKT, PTEN, and TGF-β are shown to be inhibited by different miRNAs, which in turn inhibit collagen synthesis.
It is an interesting fact that miR-668 and miR-137 were upregulated during organismal aging, which can also promote the aging
of keratinocytes in humans [65]. The study conducted by Rivetti et al. finds
that the upregulated miR-181 expression, miR-130, and miR-138 target sirtuin 1 (Sirt 1) and p63 mRNAs
[66, 67]. It is suggested that sirt1 activity
is important for replicative senescence of keratinocytes. On the other hand, p63 is actively involved in aging
[66, 68]. Additionally, miR-191 inhibits the
G1-S phase transition, which results in cell cycle arrest. This quiescent stage contributes to the aging process. MiR-152 can
drastically reduce dermal fibroblast adhesion by suppressing integrin alpha 5, a signaling protein involved in cell
surface-mediated processes [13]. Moreover, activation of p53, MAPK, and several other
signaling pathways is caused by decreased levels of miR-106 and miR-17 in aged dermal fibroblasts
[69]. Taken together, specific miRNAs have the ability to hinder the normal aging of
cells by promoting cell proliferation. The expression of miRNA is modified when the skin is exposed to UV light for a long time,
the levels of miR-27a, miR-145, miR-383, and miR-1246 are elevated, while those of miR-155, miR-663b, miR-3648, and miR-6879 are
reduced [70]. Meanwhile, UVA radiation downregulates miR-155, which causes c-Jun to be
upregulated. This influences the activity of the collagen gene in human fibroblasts [71].
Premature aging is the main result of photoaging. Human dermal fibroblasts are more sensitive to solar radiation than epidermal
keratinocytes.
Conditions like diabetic foot ulcers, pressure ulcers, and venous leg ulcers contribute to skin aging by causing chronic
inflammation, tissue damage, and impaired healing processes. These ulcers lead to persistent open wounds that disrupt the skin's
barrier function, making it susceptible to further damage and infection. The chronic inflammation associated with these conditions
accelerates the breakdown of collagen and elastin which maintain skin structure and elasticity. This will significantly impact
overall skin health [72]. Research on miRNAs highlights their critical role in skin
aging and diseases like chronic wound healing in the elderly. Key miRNAs such as miR-21, miR-31, and miR-203 promote keratinocyte
functions essential for wound healing, while miR-29b, miR-98, and miR-185 regulate collagen production and scar formation. These
findings suggest that miRNA-based therapies could enhance chronic wound healing and reduce scarring, making them valuable in
managing skin aging-related conditions [73]. Some miRNAs and their roles in dermal
aging are shown in Table 1. Meanwhile, the therapeutic potential of miRNAs in skin
aging is an evolving and active area of research today.
Table 1
miRNAs that affect different skin cells which leads to skin aging.
| miRNAs | Cell types affected | Up/Down regulated | Species (Human/Mouse) | Reference |
|---|---|---|---|---|
| miR-124 | UV radiation induces this in keratinocytes | Upregulated | Human | [74] |
| miR-23a | Keratinocytes and fibroblasts | Upregulated | Human | [73] |
| miR-779 | Langerhans cells | Upregulated | Mouse | [11] |
| miR-365 | Fibroblasts | Upregulated | Human | [11] |
| miR-124 | Keratinocytes | Upregulated | Human | [74] |
| miR-142 | Melanocytes | Altered | Human | [74] |
| miR-9 | Langerhans | Upregulated | Mouse | [11] |
| miR-25 | Melanocytes | Upregulated | Human | [11] |
| miR-151a-5p | Langerhans cells | Upregulated | Mouse | [75] |
| miR-20a | Langerhans cells | Upregulated | Mouse | [11] |
| miR-106a | Fibroblast | Downregulated | Human | [11, 76] |
| miR-148a | Primary human fibroblast | Upregulated | Human | [76, 77] |
| miR-574-3p | Senescent fibroblast | Upregulated | Human | [11] |
| miR-93 | Epidermis | Upregulated | Human | [11] |
| miR-146a | Fibroblast | Downregulated | Human | [13, 76] |
| miR-191 | Keratinocytes | Upregulated | Human | [13, 78] |
| miR-29a | Fibroblasts | Upregulated | Human | [13, 79] |
| miR-181a | Keratinocytes/fibroblasts | Upregulated | Human | [72] |
| miR-155 | Fibroblast | Downregulated | Human | [67, 80] |
miRNAs play a crucial role in the aging characteristics of various skin appendages, such as hair follicles, sebaceous glands,
and sweat glands other than above mentioned cells. Hair follicles containing hair papilla, matrix, root sheath, and bulge, have
distinct compositions. miR-191 is involved in keratinocytes senescence in hair follicles by regulating CDK6 and SATB1, thereby
influencing cell cycle progression and chromatin remodeling [11,
81]. Sebaceous glands, essential for skin lubrication and protection, are influenced by
key signaling pathways such as Wnt, c-Myc, etc., and their development is closely linked to hair follicle stem cells.
Aging affects the sebaceous gland through changes in miRNA expression, like miR-338-3p and miR-574-3p, which impact lipid synthesis
and inflammation, leading to altered skin barrier function [11,
82]. Similarly, sweat glands—the appendages for thermoregulation and fluid balance—are
categorized into eccrine and apocrine glands. These glands, which are widely distributed except in specific areas, consist of a
coiled secretory portion and a duct. The development and regeneration of sweat gland involve signaling pathways, including ERK,
Wnt, etc., which regulate cell growth and differentiation. Aging impacts sweat gland function through changes in miRNA
expression, such as miR-29 and miR-141, which affect sweat production and require further research to understand and mitigate the
age-related decline in sweat gland activity [11,
82, 83]. Understanding these miRNAs related to
aging processes may help to develop treatments to maintain skin health and combat aging.
Therapeutic interventions in anti-aging are aimed at preventing or slowing aging or the effects of aging. Beyond aesthetic
concerns, skin aging is linked to several factors of physical health and overall quality of life. Therefore, anti-aging therapy
is essential and contributes to a high quality of life. Many miRNAs have become universal regulators of a variety of skin
conditions and can function as potential therapeutic targets and novel biomarkers of disease etiology and pathogenesis
[2]. MiR-29, miR-124, and miR-152 are involved in the generation and regulation of
collagen synthesis. Numerous studies have shown that increased levels of miR-29 inhibitors increase elastin mRNA and protein
levels in vitro or in vivo. Several studies have shown that miR-29 increases the concentration of MMPs and
thereby decreases collagen synthesis. Hence, increasing the expression of collagen, fibrillin, and elastin in skin fibroblasts
results from suppressing miR-29 with an miR-29 antagonist. Such research may lead to the development of very potent cosmetic
treatments to stop the aging process of the skin [84,
85]. Therefore, modulation of their function could influence the ability of the skin to
maintain its integrity and strength. Thus, they can be considered as novel biomarkers and therapeutic targets. miRNAs have a
greater significance in anti-aging mechanisms as they are essential for the molecular regulation of several aging-related
processes [86, 87]. A study by Wing-fu
Lai et al. describes the evidence of age-associated changes in miRNA expression, which highlights the potential of miRNA
as a target and tool for anti-aging therapy [88]. Further research on miRNA finds that
miR-101 helps modulate senescence induced by UV-B radiation, and another research shows that miR-146a inhibits UV-induced aging
by targeting smad-4 [73]. Anti-aging treatment is an ongoing research area and miRNA is
of great importance in it. Silencing gene expression by using miRNAs is one method to reduce skin aging.
Figure 6 shows a miRNAbased method for skin aging therapy. Modification of miRNA
expression is emerging as a valuable focus in the cosmetic area, offering potential for the development of skin care
products [73, 89]. Current research is
shedding light on the molecular mechanisms behind several aspects of aging, including the connection between some aging
processes and individual gene activity. Thus, by inhibiting these undesirable gene activities, miRNAs may be utilized
to create new cosmetic designs and products, especially for skin care purposes like anti-aging. The expression of
tyrosinase, an enzyme essential for melanin synthesis, can be reduced using miRNAs. This technique is a new and practical method
for skin whitening [84, 90,
91]. Research on miRNA and anti-aging is still ongoing, and a deeper knowledge of the
regulatory network is needed to develop targeted and efficient therapies.
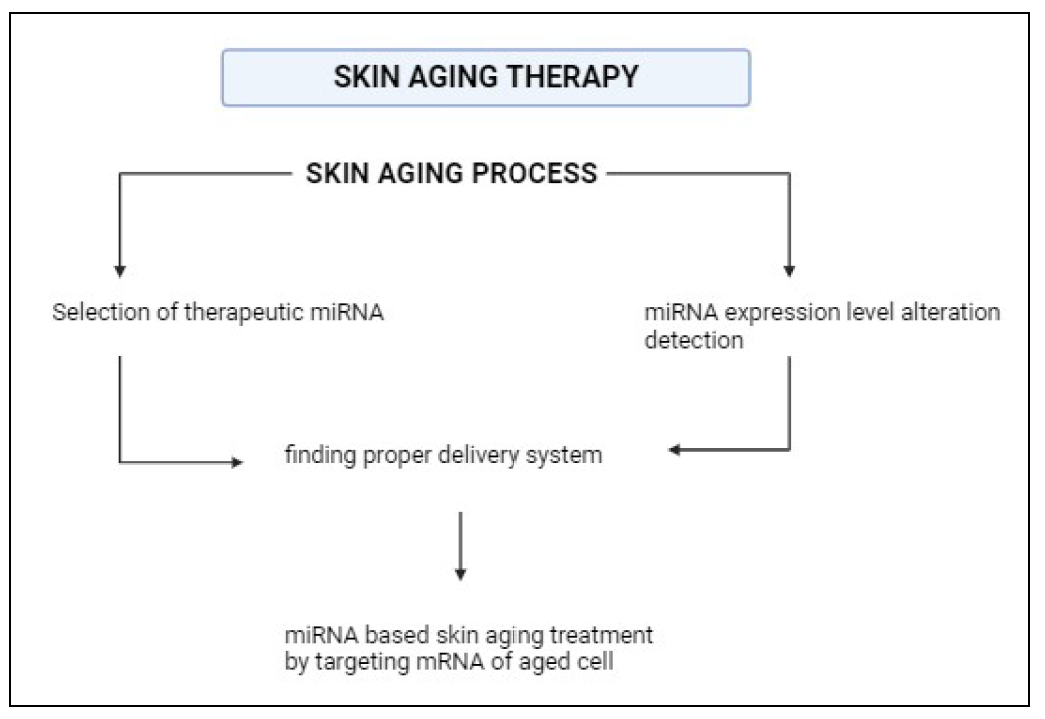
Figure 6. miRNA-based therapy. Identification of miRNAs whose expression levels are altered during aging. A suitable therapeutic miRNA is also selected. Then, using a suitable delivery system, the miRNA is transferred to the aged cell by targeting the mRNA.
Conclusions
Skin aging is a complex and integral phenomenon. It consists of 2 types: intrinsic and photoaging (extrinsic). Intrinsic aging is primarily caused by genetic factors, while extrinsic aging is caused by environmental factors. Intrinsically aged skin shows smooth, thin, sagging, pale morphology, while extrinsic aging mainly affects exposed areas, such as the neck, face, etc. Deep wrinkles, changes in pigmentation, roughness, etc. are the main features of photoaging. The commonly known skin-aging mechanism is the degradation of collagen by MMPs due to the accumulation of ROS. Skin aging is characterized by interconnected processes at the cellular, molecular, and organ levels. Despite ongoing research, the identification of distinctive biomarkers for aging remains a topic of exploration. In this review, we focus on the role of miRNA in skin aging. We have explained the current knowledge on miRNA's roles in molecular pathways and an insight into the therapeutic functions. MiRNAs play an important role in cellular and molecular functions of the skin due to their ability to control the expression of genes after transcription. Their complex regulatory roles in skin aging and dermatology make them attractive biomolecules for further study and potential therapeutic intervention. However, the main problem or challenge in this therapeutic intervention today is the knowledge of the molecular mechanisms of aging. Further research on miRNAs linked to skin aging will include the discovery of new targets and their roles. Moreover, it will provide information on the regulation of aging mechanisms at the cellular and molecular levels. Investigating miRNAs as biomarkers of skin aging and exploring their role in diagnosis and prognosis is the main future scope of this study. By focusing on these perspectives for future research, researchers can expand their knowledge in this area. This, in turn, may open avenues for effective therapeutic interventions to maintain skin health and reduce the effects of aging.
Declarations
Author contributions
Ashikha Shirin Usman PP collected articles and wrote manuscripts. Durairaj Sekar initiated the study, revised, and finalized the manuscript.
Availability of data and materials
Not applicable.
Financial support and sponsorship
None.
Conflicts of interest
All authors declared that there is no conflict of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
References
1. Horsburgh S, Fullard N, Roger M, Degnan A, Todryk S, Przyborski S, et al. MicroRNAs in the skin: role in development, homoeostasis and regeneration. Clin Sci (Lond), 2017, 131(15): 1923-1940. [Crossref]
2. Singhvi G, Manchanda P, Krishna Rapalli V, Kumar Dubey S, Gupta G, & Dua K. MicroRNAs as biological regulators in skin disorders. Biomed Pharmacother, 2018, 108: 996- 1004. [Crossref]
3. Rahmouni M, Laville V, Spadoni JL, Jdid R, Eckhart L, Gruber F, et al. Identification of new biological pathways involved in skin aging from the analysis of French women genome-wide data. Front Genet, 2022, 13: 836581. [Crossref]
4. Csekes E, & Račková L. Skin aging, cellular senescence and natural polyphenols. Int J Mol Sci, 2021, 22(23): 12641. [Crossref]
5. Farage MA, Miller KW, Elsner P, & Maibach HI. Characteristics of the aging skin. Adv Wound Care (New Rochelle), 2013, 2(1): 5-10. [Crossref]
6. Russell-Goldman E, & Murphy GF. The pathobiology of skin aging: new insights into an old dilemma. Am J Pathol, 2020, 190(7): 1356-1369. [Crossref]
7. Smith-Vikos T, & Slack FJ. MicroRNAs and their roles in aging. J Cell Sci, 2012, 125(Pt 1): 7-17. [Crossref]
8. Ruddy EK. The effect of intrinsic and extrinsic influences on skin ageing within associated demographics. Newcastle University, 2022.
9. Wang AS, & Dreesen O. Biomarkers of cellular senescence and skin aging. Front Genet, 2018, 9: 247-256. [Crossref]
10. Sotiropoulou PA, & Blanpain C. Development and homeostasis of the skin epidermis. Cold Spring Harb Perspect Biol, 2012, 4(7): a008383. [Crossref]
11. Gerasymchuk M, Cherkasova V, Kovalchuk O, & Kovalchuk I. The role of microRNAs in organismal and skin aging. Int J Mol Sci, 2020, 21(15): 5281-5293. [Crossref]
12. Thum T. Non-coding RNAs in ageing. In., vol. 17, 2014: 1-2.
13. Mancini M, Lena AM, Saintigny G, Mahé C, Di Daniele N, Melino G, et al. MicroRNAs in human skin ageing. Ageing Res Rev, 2014, 17: 9-15. [Crossref]
14. Reddy CSS, PP ASU, Ganapathy DM, Ameya K, & Sekar D. MicroRNA-21 as a biomarker in terminal stage oral squamous cell carcinoma (OSCC) in the South Indian population. Oral Oncology Reports, 2024, 9: 100139. [Crossref]
15. Lopez-Ojeda W, Pandey A, Alhajj M, & Oakley AM: Anatomy, skin (integument). In: StatPearls [Internet]. 2022, edn. StatPearls Publishing.
16. Nguyen AV, & Soulika AM. The dynamics of the skin's immune system. Int J Mol Sci, 2019, 20(8): 1811-1823. [Crossref]
17. Abdo JM, Sopko NA, & Milner SM. The applied anatomy of human skin: a model for regeneration. Wound Medicine, 2020, 28: 100179. [Crossref]
18. Nowogrodski C, Damatov Y, Sapru S, & Shoseyov O. In situ synthesis of keratin and melanin chromophoric submicron particles. ACS Omega, 2023, 8(30): 26762-26774. [Crossref]
19. Lee H, Hong Y, & Kim M. Structural and functional changes and possible molecular mechanisms in aged skin. Int J Mol Sci, 2021, 22(22): 12489. [Crossref]
20. Biniek K, Kaczvinsky J, Matts P, & Dauskardt RH. Understanding age-induced alterations to the biomechanical barrier function of human stratum corneum. J Dermatol Sci, 2015, 80(2): 94-101. [Crossref]
21. Pfisterer K, Shaw LE, Symmank D, & Weninger W. The extracellular matrix in skin inflammation and infection. Front Cell Dev Biol, 2021, 9: 682414. [Crossref]
22. Brown T, & Krishnamurthy K. Histology, dermis. Treasure Island, 2020.
23. Ergin K, & Çetinkaya R. Regulation of microRNAs. miRNomics, 2022: 1-32. [Crossref]
24. Kierszenbaum AL, & Tres L. Histology and cell biology: an introduction to pathology E-Book: Elsevier Health Sciences, 2015.
25. Khalid KA, Nawi AFM, Zulkifli N, Barkat MA, & Hadi H. Aging and wound healing of the skin: A review of clinical and pathophysiological hallmarks. Life, 2022, 12(12): 2142-2154. [Crossref]
26. Fore J. A review of skin and the effects of aging on skin structure and function. Ostomy Wound Manage, 2006, 52(9): 24-35; quiz 36-27.
27. Huang HC, Chang TM, Chang YJ, & Wen HY. UVB irradiation regulates ERK1/2- and p53-dependent thrombomodulin expression in human keratinocytes. PLoS One, 2013, 8(7): e67632. [Crossref]
28. Boismal F, Serror K, Dobos G, Zuelgaray E, Bensussan A, & Michel L. Vieillissement cutané-Physiopathologie et thérapies innovantes. médecine/sciences, 2020, 36(12): 1163-1172. [Crossref]
29. Zhang S, & Duan E. Fighting against skin aging: the way from bench to bedside. Cell transplantation, 2018, 27(5): 729-738. [Crossref]
30. Puizina-Ivic N. Skin aging. Acta Dermatovenerologica Alpina Panonica Et Adriatica, 2008, 17(2): 47-58.
31. George J, Sneed K, & Pathak Y. The skin aging process and anti-aging strategies. Biomed J Sci Tech Res, 2022, 42: 33377-33386. [Crossref]
32. Aleemardani M, Trikić MZ, Green NH, & Claeyssens F. The importance of mimicking dermal-epidermal junction for skin tissue engineering: a review. Bioengineering (Basel), 2021, 8(11):148-156. [Crossref]
33. Karim PL, & Aryani IA. Anatomy and histologic of intrinsic aging skin. Bioscientia Medicina, 2021, 5(11): 1065- 1077.
34. Rybtsova N, Berezina T, Kagansky A, & Rybtsov S. Can blood-circulating factors unveil and delay your biological aging? Biomedicines, 2020, 8(12): 615-624. [Crossref]
35. Libertini G, Shubernetskaya O, Corbi G, & Ferrara N. Is evidence supporting the subtelomere-telomere theory of aging? Biochemistry (Mosc), 2021, 86(12): 1526-1539. [Crossref]
36. Vierkötter A, & Krutmann J. Environmental influences on skin aging and ethnic-specific manifestations. Dermatoendocrinol, 2012, 4(3): 227-231. [Crossref]
37. Bocheva G, Slominski RM, & Slominski AT. Neuroendocrine aspects of skin aging. Int J Mol Sci, 2019, 20(11): 2798-2805. [Crossref]
38. Clatici VG, Racoceanu D, Dalle C, Voicu C, Tomas-Aragones L, Marron SE, et al. Perceived age and life style. The specific contributions of seven factors involved in health and beauty. Maedica (Bucur), 2017, 12(3): 191- 201.
39. Umar SA, & Tasduq SA. Integrating DNA damage response and autophagy signalling axis in ultraviolet-B induced skin photo-damage: a positive association in protecting cells against genotoxic stress. RSC Adv, 2020, 10(60): 36317-36336. [Crossref]
40. Stoykova ID, Koycheva IK, Binev BK, Mihaylova LV, & Georgiev MI. Molecular approaches to prevent UV-induced premature skin aging: focus on phytochemicals as photo-protectants. Phytochemistry Reviews, 2024: 1-32. [Crossref]
41. Toutfaire M, Bauwens E, & Debacq-Chainiaux F. The impact of cellular senescence in skin ageing: a notion of mosaic and therapeutic strategies. Biochem Pharmacol, 2017, 142: 1-12. [Crossref]
42. Han A, Chien AL, & Kang S. Photoaging. Dermatologic clinics, 2014, 32(3): 291-299. [Crossref]
43. Blume-Peytavi U, Kottner J, Sterry W, Hodin MW, Griffiths TW, Watson RE, et al. Age-associated skin conditions and diseases: current perspectives and future options. Gerontologist, 2016, 56 Suppl 2: S230-242. [Crossref]
44. Berman AE, Leontieva OV, Natarajan V, McCubrey JA, Demidenko ZN, & Nikiforov MA. Recent progress in genetics of aging, senescence and longevity: focusing on cancer-related genes. Oncotarget, 2012, 3(12): 1522- 1532. [Crossref]
45. Makrantonaki E, & Zouboulis CC. William J. Cunliffe Scientific Awards. Characteristics and pathomechanisms of endogenously aged skin. Dermatology, 2007, 214(4): 352-360. [Crossref]
46. Kang SM, Han S, Oh JH, Lee YM, Park CH, Shin CY, et al. A synthetic peptide blocking TRPV1 activation inhibits UV-induced skin responses. J Dermatol Sci, 2017, 88(1): 126-133. [Crossref]
47. Rittié L, & Fisher GJ. Natural and sun-induced aging of human skin. Cold Spring Harb Perspect Med, 2015, 5(1): a015370. [Crossref]
48. Batista LFZ, Kaina B, Meneghini R, & Menck CFM. How DNA lesions are turned into powerful killing structures: insights from UV-induced apoptosis. Mutat Res, 2009, 681(2-3): 197-208. [Crossref]
49. Sun J, Zhao H, Shen C, Li S, Zhang W, Ma J, et al. Tideglusib promotes wound healing in aged skin by activating PI3K/Akt pathway. Stem Cell Res Ther, 2022, 13(1): 269- 279. [Crossref]
50. Marinho CG, Mermejo LM, Salvatori R, Assirati JAJ, Oliveira CRP, Santos EG, et al. Occurrence of neoplasms in individuals with congenital, severe GH deficiency from the Itabaianinha kindred. Growth Horm IGF Res, 2018, 41: 71-74. [Crossref]
51. Barros-Oliveira CS, de Jesus MJM, Campos VC, Salvatori R, de Souza Araújo AA, Neto RFS, et al. Skin assessment in congenital untreated isolated GH deficiency. Endocrine, 2024. [Crossref]
52. Saccon TD, Schneider A, Marinho CG, Nunes ADC, Noureddine S, Dhahbi J, et al. Circulating microRNA profile in humans and mice with congenital GH deficiency. Aging Cell, 2021, 20(7): e13420. [Crossref]
53. Barros-Oliveira CS, Salvatori R, Dos Santos JSS, Santos PFC, Oliveira-Santos AA, Marinho CG, et al. Sweat and vitamin D status in congenital, lifetime, untreated GH deficiency. Endocrine, 2019, 65(3): 710-713. [Crossref]
54. Zhai Z, Mu T, Zhao L, Li Y, Zhu D, & Pan Y. MiR-181a-5p facilitates proliferation, invasion, and glycolysis of breast cancer through NDRG2-mediated activation of PTEN/ AKT pathway. Bioengineered, 2022, 13(1): 83-95. [Crossref]
55. Kinser HE, & Pincus Z. MicroRNAs as modulators of longevity and the aging process. Hum Genet, 2020, 139(3): 291-308. [Crossref]
56. Nikolajevic J, Ariaee N, Liew A, Abbasnia S, Fazeli B, & Sabovic M. The role of microRNAs in endothelial cell senescence. Cells, 2022, 11(7): 1185-1195. [Crossref]
57. Fisher GJ, Wang B, Cui Y, Shi M, Zhao Y, Quan T, et al. Skin aging from the perspective of dermal fibroblasts: the interplay between the adaptation to the extracellular matrix microenvironment and cell autonomous processes. J Cell Commun Signal, 2023, 17(3): 523-529. [Crossref]
58. Berezikov E, Guryev V, van de Belt J, Wienholds E, Plasterk RH, & Cuppen E. Phylogenetic shadowing and computational identification of human microRNA genes. Cell, 2005, 120(1): 21-24. [Crossref]
59. Dexheimer PJ, & Cochella L. MicroRNAs: from mechanism to organism. Front Cell Dev Biol, 2020, 8: 409-419. [Crossref]
60. Baptista B, Riscado M, Queiroz JA, Pichon C, & Sousa F. Non-coding RNAs: Emerging from the discovery to therapeutic applications. Biochem Pharmacol, 2021, 189: 114469. [Crossref]
61. O'Brien J, Hayder H, Zayed Y, & Peng C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol (Lausanne), 2018, 9: 402-412. [Crossref]
62. Elder CR, & Pasquinelli AE. New roles for microRNAs in old worms. Front Aging, 2022, 3: 871226. [Crossref]
63. de Lencastre A, Pincus Z, Zhou K, Kato M, Lee SS, & Slack FJ. MicroRNAs both promote and antagonize longevity in C. elegans. Curr Biol, 2010, 20(24): 2159-2168. [Crossref]
64. He J, & Jiang BH. Interplay between reactive oxygen species and microRNAs in cancer. Curr Pharmacol Rep, 2016, 2(2): 82-90. [Crossref]
65. Potter ML, Hill WD, Isales CM, Hamrick MW, & Fulzele S. MicroRNAs are critical regulators of senescence and aging in mesenchymal stem cells. Bone, 2021, 142: 115679. [Crossref]
66. Cong L, Ma J, Zhang Y, Zhou Y, Cong X, & Hao M. Effect of anti-skin disorders of ginsenosides- a systematic review. J Ginseng Res, 2023, 47(5): 605-614. [Crossref]
67. Rivetti di Val Cervo P, Lena AM, Nicoloso M, Rossi S, Mancini M, Zhou H, et al. p63-microRNA feedback in keratinocyte senescence. Proc Natl Acad Sci USA, 2012, 109(4): 1133-1138. [Crossref]
68. Chen Z, Shentu TP, Wen L, Johnson DA, & Shyy JY. Regulation of SIRT1 by oxidative stress-responsive miRNAs and a systematic approach to identify its role in the endothelium. Antioxid Redox Signal, 2013, 19(13): 1522-1538. [Crossref]
69. Dong Y, Chen H, Gao J, Liu Y, Li J, & Wang J. Bioactive ingredients in Chinese herbal medicines that target noncoding RNAs: promising new choices for disease treatment. Front Pharmacol, 2019, 10: 515-525. [Crossref]
70. Zhang Y, Yang C, Yang S, & Guo Z. MiRNA-27a decreases ultraviolet B irradiation-induced cell damage. J Cell Biochem, 2020, 121(2): 1032-1038. [Crossref]
71. Song J, Liu P, Yang Z, Li L, Su H, Lu N, et al. MiR-155 negatively regulates c-Jun expression at the post-transcriptional level in human dermal fibroblasts in vitro: implications in UVA irradiation-induced photoaging. Cell Physiol Biochem, 2012, 29(3-4): 331-340. [Crossref]
72. Xie J, Wu W, Zheng L, Lin X, Tai Y, Wang Y, et al. Roles of microRNA-21 in skin wound healing: a comprehensive review. Front Pharmacol, 2022, 13: 828627. [Crossref]
73. Li X, Ponandai-Srinivasan S, Nandakumar KS, Fabre S, Xu Landén N, Mavon A, et al. Targeting microRNA for improved skin health. Health Sci Rep, 2021, 4(4): e374. [Crossref]
74. Harada M, Jinnin M, Wang Z, Hirano A, Tomizawa Y, Kira T, et al. The expression of miR-124 increases in aged skin to cause cell senescence and it decreases in squamous cell carcinoma. Biosci Trends, 2017, 10(6): 454-459. [Crossref]
75. Noren Hooten N, Fitzpatrick M, Wood WH, 3rd, De S, Ejiogu N, Zhang Y, et al. Age-related changes in microRNA levels in serum. Aging (Albany NY), 2013, 5(10): 725- 740. [Crossref]
76. Stratz C, Nührenberg TG, Binder H, Valina CM, Trenk D, Hochholzer W, et al. Micro-array profiling exhibits remarkable intra-individual stability of human platelet micro-RNA. Thromb Haemost, 2012, 107(4): 634-641. [Crossref]
77. ElSharawy A, Keller A, Flachsbart F, Wendschlag A, Jacobs G, Kefer N, et al. Genome-wide miRNA signatures of human longevity. Aging Cell, 2012, 11(4): 607-616. [Crossref]
78. Lena AM, Mancini M, Rivetti di Val Cervo P, Saintigny G, Mahé C, Melino G, et al. MicroRNA-191 triggers keratinocytes senescence by SATB1 and CDK6 downregulation. Biochem Biophys Res Commun, 2012, 423(3): 509-514. [Crossref]
79. Martinez I, Cazalla D, Almstead LL, Steitz JA, & DiMaio D. miR-29 and miR-30 regulate B-Myb expression during cellular senescence. Proc Natl Acad Sci U S A, 2011, 108(2): 522-527. [Crossref]
80. Mancini M, Lena AM, Saintigny G, Mahé C, Di Daniele N, Melino G, et al. MicroRNAs in human skin ageing. Ageing research reviews, 2014, 17: 9-15. [Crossref]
81. Yadav S, Sarkar S, Pandey A, & Singh T. Ageing at molecular level: role of microRNAs. Subcell Biochem, 2023, 102: 195-248. [Crossref]
82. Weng T, Wu P, Zhang W, Zheng Y, Li Q, Jin R, et al. Regeneration of skin appendages and nerves: current status and further challenges. J Transl Med, 2020, 18(1): 53-64. [Crossref]
83. Zonnefeld AG, Cui CY, Tsitsipatis D, Piao Y, Fan J, MazanMamczarz K, et al. Characterization of age-associated gene expression changes in mouse sweat glands. Aging (Albany NY), 2024, 16(8): 6717-6730. [Crossref]
84. Zhang P, Chen J, Li T, & Zhu YY. Use of small RNA as antiaging cosmeceuticals. J Cosmet Sci, 2013, 64(6): 455- 468.
85. Harmanci D, Erkan EP, Kocak A, & Akdogan GG. Role of the microRNA-29 family in fibrotic skin diseases. Biomed Rep, 2017, 6(6): 599-604. [Crossref]
86. Lehmann TP, Guderska U, Kałek K, Marzec M, Urbanek A, Czernikiewicz A, et al. The regulation of collagen processing by miRNAs in disease and possible implications for bone turnover. Int J Mol Sci, 2021, 23(1): 91-104. [Crossref]
87. Condrat CE, Thompson DC, Barbu MG, Bugnar OL, Boboc A, Cretoiu D, et al. MiRNAs as biomarkers in disease: latest findings regarding their role in diagnosis and prognosis. Cells, 2020, 9(2): 276-289. [Crossref]
88. Lai WF, Lin M, & Wong WT. Tackling aging by using miRNA as a target and a tool. Trends Mol Med, 2019, 25(8): 673-684. [Crossref]
89. Lawrence P, & Ceccoli J. Advances in the application and impact of microRNAs as therapies for skin disease. BioDrugs, 2017, 31(5): 423-438. [Crossref]
90. Chen JS, & Wu DT. Application of intronic microRNA agents in cosmetics. Methods Mol Biol, 2013, 936: 325- 341. [Crossref]
91. Eshkoor SA, Ghodsian N, & Akhtari-Zavare M. MicroRNAs influence and longevity. Egyptian Journal of Medical Human Genetics, 2022, 23(1): 105-115. [Crossref]